Les origines de la vie (André Brack)
Texte extrait du GDR
Exobiologie
(http://www.exobio.cnrs.fr/article.php3?id_article=23)
Dans toutes les civilisations, l'homme a été intrigué par son
origine et par l'origine de la vie. Pendant des millénaires, la théorie
confortable et rassurante de la "génération spontanée" répondit à
cette interrogation. Déjà dans la Chine ancienne on pensait que les bambous
donnaient spontanément naissance aux pucerons. Les écrits sacrés de l'Inde
mentionnent la naissance spontanée de mouches à partir d'ordures et de sueur.
Les inscriptions babyloniennes signalent que la boue des canaux pouvait
engendrer des vers. Dans l'Egypte antique, on pensait que le limon déposé par
le Nil pouvait générer spontanément grenouilles et crapauds. Bien que partant
d'observations réelles - la présence d'animaux dans différents milieux - ces
civilisations furent incapables d'en dégager une interprétation correcte, faute
de pouvoir recourir à la vérification expérimentale.
Pour les philosophes grecs, la vie est propriété même de la
matière ; elle est éternelle et apparait
spontanément chaque fois que les conditions sont propices. Ces idées
apparaissent clairement dans les écrits de Thalès, de Démocrite, d'Epicure, de
Lucrèce et même dans ceux de Platon. Aristote réalise la synthèse des idées
précédemment émises. Avec lui, la génération spontanée acquiert la dimension
d'une véritable théorie. Celle-ci traverse allègrement le Moyen-Age
et la Renaissance ; de grands penseurs comme Newton, Descartes et Bacon la
soutiennent. C'est à cette époque qu'apparaissent les premières
expérimentations sur la génération spontanée. C'est ainsi qu'au milieu du 17e
siècle, Van Helmont, un médecin flamand déclara obtenir des souris avec des
grains de blé et une chemise imprégnée de sueur humaine. Van Helmont constate
avec surprise que les souris obtenues dans ces conditions sont identiques à
celles nées par procréation.
Observation et expérimentation approximative renforcent l'idée
fausse au lieu de la remettre en cause. Les premières attaques expérimentales
furent portées par Rédi, un médecin naturaliste
toscan. Dans son traité de 1668, il publie une série d'expériences démontrant
que les asticots ne sont pas générés spontanément par la viande en putréfaction
lorsque l'on prend la précaution de recouvrir les bocaux d'une très fine
mousseline. Six ans après la parution du traité de Rédi,
le savant hollandais Anton Van Leeuwenhoek effectue les premières observations
de microorganismes à travers un microscope de sa fabrication. Dès lors, on
trouve des microorganismes partout et les adeptes de la génération spontanée se
réfugient dans le monde de l'infiniment petit. Cependant, Van Leeuwenhoek
pensait déjà que ces microorganismes provenaient d'une contamination des
solutions par l'air ambiant. En 1718, son disciple Louis Joblot
démontre expérimentalement que les microorganismes proviennent bien de l'air
ambiant. Il ne réussit pas à convaincre les naturalistes qui considéraient le
monde des microorganismes comme le bastion le plus significatif de la
génération spontanée. Il est vrai que, depuis Rédi,
on cessa progressivement de croire à l'intervention de la génération spontanée
dans le cas des êtres vivants hautement organisés. Même Buffon, au milieu du
18e siècle, pense que la nature est pleine de germes de vie capables de
s'éparpiller lors du pourissement puis de s'unir pour
produire des microbes. John Needham, l'ami gallois de
Buffon, réalise de nombreuses expériences pour confirmer cette conception. Il
place différentes substances organiques dans une fiole hermétiquement close et
chauffe l'ensemble pour stériliser. Après traitement, toutes les solutions
foisonnent de microbes. L'abbé italien Lazzaro
Spallanzani porte les solutions à des températures plus élevées et contredit Needham. Une vive polémique s'engagea sur l'effet de la
température mais Spallanzani n'arriva pas à imposer ses vues. Une fois encore,
la notion subjective de la "force vitale" l'emporta sur la
démonstration expérimentale.
Cette controverse va atteindre son apothéose un siècle plus tard
avec la publication en 1860 du traité de Félix Pouchet " Hétérogénie ou
traité de la génération spontanée". Dans son ouvrage, Félix Pouchet,
correspondant de l'Institut, définit la génération spontanée comme étant la
production d'un être organisé nouveau, dénué de parents, et dont tous les
éléments primordiaux ont été tirés de la matière ambiante. Louis Pasteur se
livre alors à des expériences rigoureuses sur le pouvoir fermentescible et la
stabilité à la chaleur des germes de l'air ambiant et
réfute tous les arguments de Pouchet. Pour arbitrer le conflit, l'Académie des
Sciences charge une Commision de faire reproduire les
expériences devant ses membres. Le 22 juin 1864, Louis Pasteur démontre que ce
sont les germes de l'air ambiant qui altèrent les solutions aqueuses, étayant
ainsi la théorie des germes. Le 20 février 1865, la Commission de l'Académie
des Sciences donnait raison à Louis Pasteur contre Félix Pouchet : la
matière ambiante ne peut pas générer un être organisé nouveau, dénué de
parents. Elle mettait fin à une idée fausse qui dû probablement sa longévité au
poids culturel de la notion de vie. Pasteur eut le désir de créer la vie, mais
probablement une vie obtenue par synthèse chimique à partir de molécules
simples. En 1878 il écrivait : « La génération spontanée, je la
cherche sans la découvrir depuis vingt ans. Non, je ne la juge pas
impossible ». Formidable encouragement aux chimistes de l'origine de la
vie terrestre. En démontrant que la vie ne peut provenir que d'êtres vivants
préexistants, Pasteur transforme le mystère en énigme : la vie a une
histoire et donc un commencement.
Propriétés fondamentales des systèmes vivants
Des têtes sans jambes, des jambes sans têtes, des bras, des
torses, jonchaient autrefois la surface de la Terre. Un jour, par hasard, tous
les éléments nécessaires à la constitution d'un être humain se rencontrèrent et
s'auto-organisèrent. Ainsi naquit, selon Empédocle,
le premier être vivant. L'image d'Empédocle peut être transposée au niveau
moléculaire : un certain nombre de molécules s'auto-organisent
dans l'eau et forment des structures chimiques, sortes d'automates chimiques,
capables d'assembler d'autres molécules pour générer des structures à leur
image, produisant ainsi plus d'eux-mêmes par eux-mêmes (Figure 1). C'est l'auto-reproduction. Par suite de légères erreurs de montage,
des automates plus aptes à se reproduire apparaissent et deviennent les espèces
dominantes. C'est l'évolution. Auto-reproduction et
évolution sont donc les deux qualités qui caractérisent, a minima, le passage
de la matière à la vie.
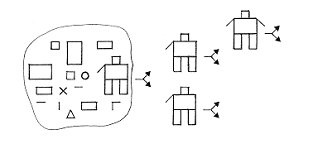
Figure 1.
Représentation schématique de petits automates chimiques capables d'auto-reproduction et d'évolution (crédit A. Brack)
Comment passer de la matière à la vie
Les scientifiques supposent que les premiers petits automates
chimiques apparurent dans l'eau des océans il y a environ 4 milliards d'années.
L'eau ne pose pas de problème majeur car la Terre était déjà couverte d'eau peu
de temps après sa formation, il y a plus de 4 milliards d'années, comme
l'indiquent les rapports des isotopes de l'oxygène mesurés dans des zircons (cristaux
de silicate de zirconium contenant des traces d'uranium et de thorium qui
permettent de les dater) vieux de 4,4 milliards d'années retrouvés dans des
sédiments d'Australie occidentale.
Dès 1953, qui marqua le début de la chimie prébiotique
avec l'expérience de Miller, les chimistes ont cherché à reconstituer dans
leurs tubes à essais des automates ressemblant à une cellule simplifiée en
raison de la remarquable unité du vivant contemporain et de son mode de
fonctionnement cellulaire. Pendant des décennies, les chimistes se sont ainsi
évertués a reconstituer en laboratoire des molécules indispensables au
fonctionnement d'une cellule : les molécules de compartimentation
(molécules membranaires), les molécules de l'information (ARN et ADN) et les
molécules catalytiques (enzymes protéiques).
La matière première des automates primitifs
Par analogie avec le monde vivant contemporain, on considère
généralement que la vie primitive utilisait déjà des molécules organiques,
échafaudages d'atomes de carbone liés à des atomes d'hydrogène, d'oxygène,
d'azote, de phosphore et de soufre. Les formes de carbone les plus simples
capables de conduire aux molécules organiques sont gazeuses : dioxyde de
carbone et monoxyde de carbone pour les formes oxydées et méthane pour la forme
réduite.
Quand on pense molécules gazeuses, on pense tout naturellement à
l'atmosphère terrestre. L'idée de composés chimiques fabriqués dans
l'atmosphère terrestre fut d'abord émise par le biochimiste russe Alexandre Oparin en 1924, puis par l'Anglais John Haldane en 1929,
indépendamment d'Oparin. Oparin
pensait que l'atmosphère primitive était dominée par le méthane, une forme
réduite du carbone alors que pour Haldane, les molécules organiques se seraient
formées à partir de dioxyde de carbone, une forme oxydée du carbone.
L'hypothèse d'Oparin se trouva confortée en 1953 par
l'expérience remarquable du jeune chimiste américain Stanley Miller. Ce dernier
remplit un ballon d'un mélange gazeux de méthane, d'ammoniac, d'hydrogène et de
vapeur d'eau et soumit ce mélange à l'action d'un arc électrique simulant les
orages de la Terre primitive. Parmi les composés formés, il identifia l'acide
cyanhydrique et le formaldéhyde, précurseurs chimiques des briques du vivant.
Il isola également plusieurs acides aminés, dont la glycine, le plus simple des
acides aminés protéiques. Depuis l'expérience de Miller, dix-sept des vingt
acides aminés protéiques ont été isolés ainsi que certains éléments
constitutifs des acides nucléiques. Cependant, les géochimistes affirment
aujourd'hui que l'atmosphère terrestre primitive était essentiellement neutre,
formée majoritairement de dioxyde de carbone, d'eau, et d'hydrogène sulfuré
d'origine volcanique ou micrométéoritique, avec des
quantités mineures d'autres gaz tels que méthane, monoxyde de carbone et azote.
Lorsque l'on refait l'expérience de Miller en passant progressivement du
méthane au dioxyde de carbone, la formation d'acides aminés devient de plus en
plus difficile. Si l'atmosphère primitive était réellement dominée par du
dioxyde de carbone, elle ne pouvait pas être la source exclusive de la matière
organique nécessaire à l'émergence de la vie terrestre et d'autres filières ont
dû contribuer à la production de pièces d'automates chimiques, filières
océanique et spatiale.
Les sources hydrothermales sous-marines présentent un
environnement favorable aux synthèses prébiotiques.
Lorsque deux plaques tectoniques s'écartent, le magma remonte et se solidifie
pour former les dorsales océaniques. Au cours de son ascension et de son
refroidissement, le magma se contracte et se fissure. L'eau de mer s'infiltre
sur plusieurs centaines de mètres de profondeur et se réchauffe au contact du
basalte chaud jusqu'à atteindre des températures de 350 °C. L'eau se charge en
gaz, hydrogène, azote, oxyde de carbone, dioxide de
carbone, méthane, anhydride sulfureux, hydrogène sulfuré, puis s'échappe du
fond de l'océan sous forme de véritables geysers. Les sources hydrothermales
sous-marines constituent un milieu exceptionnel qui a peu évolué depuis quatre
milliards d'années. Les éléments indispensables à la fabrication des pièces
d'automates chimiques y sont présents : hydrogène, azote, monoxyde et
dioxyde de carbone, hydrogène sulfuré, méthane et, bien sûr, eau. Le magma
fournit en continu l'énergie nécessaire sous forme de chaleur. Le milieu est
protégé des effets destructeurs des rayons ultraviolets par la couche d'eau
océanique qui amortit également le bombardement météoritique. Est-ce là le
berceau des automates chimiques ? Une température de 350°C est trop élevée
pour permettre la survie des automates, voire même des pièces d'automates. Le
record pour les bactéries hyperthermophiles qui
vivent à des températures supérieures à 80°C au voisinage des sources
hydrothermales est actuellement de 113°C. Il semble bien que ces bactéries hyperthermophiles proviennent de bactéries vivant à des
températures plus agréables, inférieures à 80°C, qui se seraient adaptées
relativement récemment à ces températures extrêmes. Il est donc peu probable
que les sources hydrothermales aient été le berceau
des automates chimiques mais elles ont très bien pu produire certaines des
pièces nécessaires à leur émergence. Pour l'Allemand Günter Wächtershäuser,
les molécules organiques primordiales se seraient formées au niveau de ces
sources hydrothermales par réduction des oxydes de carbone grâce à la réaction
de l'hydrogène sulfuré sur le sulfure de fer. En laboratoire, le sulfure de
fer, l'hydrogène sulfuré et le dioxyde de carbone réagissent en milieu
anaérobie pour donner de l'hydrogène et une variété de dérivés soufrés. Dans
certaines conditions, il se forme des dérivés soufrés particulièrement
intéressants, qui ont peut-être constitué, selon le biologiste belge Christian
De Duve, le principal moteur chimique d'un monde
vivant primitif soufré.
Des molécules organiques d'origine extraterrestre
L'apport de molécules organiques extraterrestres reste de loin le
plus important. Les sondes Véga 1 et 2, Giotto, Suisei
et Sakigake ont montré que la comète de Halley est
riche en matériaux organiquex, le taux moyen en poids
de carbone présent dans les grains cométaires étant estimé à 14 %. Parmi les
molécules identifiées, on retrouve l'acide cyanhydrique et le formaldéhyde. Ces
composés, ainsi que de nombreuses autres molécules d'intérêt
prébiotique, ont également été observés dans les
comètes Hyakutake en 1996 et Hale-Bopp
en 1997.
Les météorites carbonées, représentées typiquement par les
météorites d'Orgueil et de Murchison, renferment des composés organiques comme
des hydrocarbures aliphatiques et aromatiques. Des hydrocarbures aromatiques
polycycliques, des kérogènes et des fullerènes sont également présents. Des
composés plus proches des composés biologiques ont été identifiés : acides
carboxyliques, acides aminés, bases nucléiques, amines, amides, alcools, etc.
La météorite carbonée de Murchison renferme plus de soixante-dix acides aminés
différents. Au nombre de ceux-ci on trouve huit acides aminés protéiques. Les
atomes de carbone qui constituent l'ossature des systèmes vivants et, très
probablement, des automates chimiques primitifs, occupent généralement le
centre de tétraèdres. Comme les groupes d'atomes liés aux quatre sommets des
tétraèdres sont différents, les atomes de carbone sont asymétriques et se
présentent sous deux formes, images l'une de l'autre dans un miroir, non
superposables. Il existe donc des acides aminés L et des acides aminés D. Les
systèmes vivants n'utilisent qu'une seule des deux formes possibles pour chaque
grande famille de constituants biologiques, protéines, ADN, etc. C'est ainsi
que les protéines n'utilisent que la forme L des acides aminés. Une vie qui
utiliserait indifféremment et simultanément les deux formes L et D des
molécules biologiques semble très improbable. Le minéralogiste américain John
Cronin trouva environ 55% de formes L et 45% de formes D pour certains acides
aminés présents dans la météorite de Murchison. La découverte par les
astronomes d'un rayonnement infrarouge fortement polarisé dans un nuage
moléculaire de la nébuleuse d'Orion pourrait expliquer l'origine extraterrestre
de cet excès de formes L.
Des collectes de poussières interplanétaires dans les glaces du Groënland et de l'Antarctique par le minéralogiste français
Michel Maurette permettent d'évaluer à environ 50 à
100 tonnes la quantité de grains interplanétaires arrivant tous les jours
actuellement à la surface de la Terre. Environ 99% de cette masse est apporté
par des micrométéorites dont le diamètre est compris entre 50 et 500 µm. Les micrométéorites sont apparentées aux météorites les
plus primitives, celles du groupe des météorites carbonées représentées par la
météorite de Murchison. Une analyse détaillée des teneurs en carbone de
différents groupes de micrométéorites permet d'estimer à cent tonnes le flux
total de carbone organique apporté tous les ans à la Terre. La quantité livrée
à la Terre pendant la phase active du bombardement terrestre entre 4,1 et 3,8
milliards d'années, quand le flux micrométéoritique
était vraisemblablement mille fois plus intense qu'aujourd'hui, est estimée à
1023g. Pour donner un ordre de grandeur, cette valeur représente cent mille
fois la valeur actuelle du carbone biologique recyclé aujourd'hui à la surface
de la Terre. Ces grains renferment une forte proportion de sulfures métalliques,
d'oxydes, d'argiles qui sont autant de catalyseurs. Au contact de l'eau
liquide, les grains ont donc pu fonctionner comme des microréacteurs chimiques transformant la matière organique des grains à l'aide des
catalyseurs présents.
Près de cent dix molécules différentes ont été identifiées à ce
jour dans les nuages denses de gaz et de poussières du milieu interstellaire.
Parmi ces molécules, quatre-vingt-trois contiennent du carbone. La présence de
glycine, longtemps controversée, a été confirmée récemment dans le milieu
interstellaire ainsi que les molécules précurseurs qui conduisent généralement
aux acides aminés : acide cyanhydrique, ammoniac et formaldéhyde. Pour
vérifier que la synthèse d'acides aminés dans les conditions du milieu
interstellaire se fait facilement, un mélange de glaces d'eau, d'ammoniac, de
méthanol, de monoxyde et de dioxyde de carbone a été irradié au Laboratoire
d'astrophysique de Leyde aux Pays-Bas, dans des conditions mimant celles du
milieu interstellaire (vide poussé de 10-7 mbar, température de -261°C). Une
fois ramenés à la température ambiante, les
échantillons, ont été analysés au Centre de biophysique moléculaire du CNRS à
Orléans. Nous y avons identifié seize acides aminés dont six (glycine, alanine,
valine, proline, serine, acide aspartique) font partie des vingt acides aminés
protéiques.
On sait que les météorites, dont la masse est supérieure à cent
grammes, peuvent transporter les acides aminés. Cependant, l'apport en
météorites (aujourd'hui de l'ordre de cent tonnes par an) est très minoritaire
par rapport à celui des micrométéorites. Il fallait donc vérifier à partir de
quelle taille, une micrométéorite devient un transporteur possible d'acides
aminés. Des expériences spatiales ont été menées en orbite terrestre pour établir
ce point. Dans l'expérience BIOPAN-1, six acides aminés L présents dans la
météorite de Murchison ont été exposés aux conditions de l'espace pendant
quinze jours en juin 1994 à bord de la capsule automatique russe FOTON-8. Ils
ont été exposés nus mais également enrobés dans une argile, la montmorillonite.
Après récupération, les acides aminés ont été analysés à l'aide d'une technique
qui permet de mesurer le taux de décomposition des acides aminés mais aussi le
taux de racémisation, c'est-à-dire la proportion de
forme L qui a été éventuellement transformée en forme D au cours de
l'exposition. Les analyses effectuées après le vol révèlent des différences
nettes de comportement se traduisant notamment par un déficit significatif des
acides aminés acides, acide glutamique et acide aspartique dans les
échantillons exposés au rayonnement solaire. Aucun déficit n'est constaté
lorsque les échantillons sont associés à des argiles. Aucune racémisation n'a été observée. Le second vol de dix jours
en 1997 a confirmé les résultats du premier vol et a permis de montrer
qu'au-dessous de 5 µm, l'argile n'offrait plus de
protection totale contre le rayonnement solaire. Un troisième vol s'est déroulé
en 1999 à bord de la station MIR. Pour cette mission, différentes protections
minérales ont été utilisées, à différentes épaisseurs : une argile, une
poudre de basalt et une poudre de météorite. Après
trois mois en orbite terrestre, les acides aminés ont été détruits à hauteur de
50% en l'absence de protection minérale. A épaisseur égale, c'est la poudre de
météorite qui a présenté le meilleur pouvoir protecteur. La poudre météoritique
protège efficacement à partir d'une épaisseur de 5 µm.
En d'autres termes, toute micrométéorite de taille supérieure à 5 µm représente un transporteur possible d'acides aminés dans
l'espace.
Les premiers automates chimiques
Comme déjà mentionné, les chimistes ont d'abord pensé que les
automates primitifs devaient ressembler à des cellules simplifiées, en
s'inspirant du vivant contemporain cellulaire et de l'universalité du code
génétique. A partir des petites molécules organiques, les chimistes se sont
efforcés de reconstituer en laboratoire les trois familles de biopolymères indispensables au fonctionnement de la
cellule. Certaines substances organiques présentes dans les météorites
carbonées de Murchison et d'Allende forment dans l'eau des cloisons qui
ressemblent à des membranes. Des acides gras sont notamment présents dans ces
météorites. Toutefois, les membranes produites à l'aide de ces composés
amphiphiles simples ne sont pas très stables, de sorte que des composés
chimiques plus complexes ont vraisemblablement été nécessaires pour conférer
une bonne stabilité aux membranes primitives.
Des mini-protéines ont été reconstituées
en laboratoire. Une équipe de chercheurs japonais, en collaboration avec notre
laboratoire à Orléans, a obtenu des chaînes renfermant jusqu'à huit acides
aminés en faisant passer alternativement une solution de glycine d'une chambre
réactionnelle à 220°C à une chambre à 0°C, mimant ainsi les conditions de
trempe thermique qui règnent à proximité des sources chaudes sous-marines. Il a
été montré par ailleurs que les argiles permettent d'obtenir de longues chaînes
de mini-protéines. Les chimistes à Orléans ont réussi
à reproduire les géométries des protéines, hélices-alpha
et feuillets-beta. Toute une série de mini-protéines construites sur une alternance stricte
d'acides aminés hydrophiles et hydrophobes a été préparée. Elles adoptent
toutes une structure en feuillets-beta dans l'eau par
agrégation des groupes hydrophobes, à condition que le caractère hydrophobe
soit bien marqué. Lorsque l'on ajoute de l'alcool à l'eau, la force des
interactions hydrophobes qui génèrent les feuillets-beta s'atténue et les mini-protéines
adoptent alors une géométrie en hélice-alpha. C'est
donc l'eau qui, par ses propriétés physiques spécifiques, permet la
structuration en feuillets-beta. Les mini-protéines à séquence alternée, capables de ce fait de
prendre la géométrie en feuillets-beta, sont très
résistantes, ce qui a peut-être permis leur sélection sur la Terre primitive.
La formation de feuillets-beta requiert l'utilisation
d'acides aminés de même chiralité, c'est-à-dire tous L ou tous D. Lorsque les
séquences alternées renferment à la fois des formes L et D distribuées au
hasard le long des chaînes, seuls les segments contenant au moins six acides
aminés L (ou D) consécutifs s'agrègent en feuillets-beta.
De nombreux mini-protéines
manifestent une activité catalytique. Par exemple, à Orléans, nous avons montré
que des mini-protéines basiques ne renfermant que
deux acides aminés différents coupent les ARN. La grande majorité des travaux
de reconstitution d'acides nucléiques prébiotiques
porte sur les ARN car ils sont considérés comme étant plus anciens, plus
primitifs, que les ADN. Certaines bases des nucléotides sont obtenues
facilement à partir de l'acide cyanhydrique ou en soumettant un mélange gazeux
de méthane, d'éthane et d'ammoniac à des décharges électriques. Les bases sont
également présentes en très petites quantités dans les météorites et peut-être
même dans les comètes. La synthèse des sucres à partir de formaldéhyde fournit
un mélange très complexe dans lequel le sucre recherché est très minoritaire.
Même si les argiles catalysent efficacement la formation des chaînes de
nucléotides, la synthèse des nucléotides, combinaison d'un phosphate, d'un
sucre et d'une base, n'a pas pu être reproduite en laboratoire dans des
conditions simples simulants l'environnement de la
Terre primitive. Les chimistes ont alors cherché à tourner la difficulté en
exploitant des analogues, succédanés des ARN. Les travaux, en cours, ont déjà
donné des résultats encourageants. Cependant, la formation prébiotique
d'ARN demeure encore inexpliquée.
Une vie primitive pré-cellulaire
Dans les années 1980, les biologistes ont découvert que certains
ARN, les ribozymes, étaient capables non seulement de véhiculer l'information
mais aussi d'exercer une activité catalytique, comme les enzymes protéiques.
Très vite s'est développé l'idée d'un monde d'ARN berceau de la vie sur Terre.
Le monde d'ARN a constitué vraisemblablement un épisode dans l'histoire de la
vie. Le fait que l'étape clé de la formation de la liaison peptidique au cours
de la biosynthèse des protéines se fasse par catalyse de l'ARN sans
intervention quelconque d'un acide aminé et la découverte récente des Mimivirus à ADN, macrovirus
possédant des gènes communs à tous les organismes des trois branches du vivant,
eucaryotes, bactéries et archea, étayent l'idée d'un
monde d'ARN ancestral. Reste maintenant à comprendre la formation prébiotique de l'ARN qui n'a trouvé, à ce jour, aucune
explication convaincante. Il est raisonnable de penser que l'émergence du monde
de l'ARN ait été préparée par des systèmes autocatalytiques
plus simples ou par des catalyseurs produisant de l'ARN en grande
quantités :
CHONS et H2O => Robots ou catalyseurs => Monde viral à ARN
=> Cellules (avec ARN, protéines et membranes)
Lorsque deux molécules A et B réagissent pour former une nouvelle
molécule A-B, il arrive quelques fois que le produit de la réaction A-B
reconnaisse et organise les éléments A et B, favorise leur couplage chimique et
catalyse ainsi sa propre formation. Ce type d'assistance chimique, appelé
autocatalyse, est courant en chimie. Cependant, les molécules A-B reproduites
par autocatalyse ne comportent que deux "lettres" car A et B ne peuvent être liés que d'un côté. De ce fait, l'information
reste qualitativement pauvre, comme serait pauvre une langue n'utilisant que
des mots à deux lettres. Comme pour l'écriture, l'emploi de molécules bifonctionnelles va permettre d'enrichir l'information à
l'infini. Se pose alors un évident problème de sélection car maintenant A et B
vont pouvoir former A-B, certes, mais également A-A, B-A et B-B. L'auto-reproduction recherchée devra donc obligatoirement
associer autocatalyse et sélection, ce qui, en dehors des ARN biologiques et de
leurs analogues, n'a pas encore été réalisé en tube à essais.
Certains scientifiques pensent que l'autocatalyse s'est développée
sur des surfaces minérales. Ils favorisent l'hypothèse selon laquelle des
systèmes chimiques utilisaient directement le dioxyde de carbone comme source
de carbone, a l'instar des plantes et de certaines bactéries. Ces organismes
vivants primitifs sont décrits comme des molécules organiques autocatalytiques se développant sur des surfaces minérales
de pyrite FeS2. Comme mentionné, la réaction du fer ferreux sur l'hydrogène
sulfureux produit d'une part de l'hydrogène qui réduit le dioxyde de carbone et
de la pyrite qui sert de support minéral à la croissance autocatalytique
du réseau organique. De ce fait, il y a croissance simultanée du réseau
organique et du support minéral. Le départ des molécules organiques de la
surface minérale au profit de molécules mieux adaptées à la surface permet au
système d'évoluer en se perfectionnant. L'idée d'une participation active des
surfaces minérales, émise dès 1953 par l'Anglais J. Desmond
Bernal, connait depuis peu
un regain d'intérêt et de nombreux laboratoires se sont lancés dans cette
chimie on the rocks.
A la recherche d'automates fossilisés
Les roches les plus anciennes susceptibles de présenter des traces
de vie sont des sédiments vieux de quelque 3,75 milliards d'années découverts
dans le sud-ouest du Groënland. Ces sédiments
témoignent de la présence permanente d'eau liquide, de gaz carbonique dans
l'atmosphère et renferment des kérogènes, molécules organiques complexes dont
les atomes de carbone sont enrichis en isotope 12. D'une manière générale, les
molécules biologiques produites par photosynthèse sont caractérisées par un
enrichissement en 12C par rapport aux carbonates minéraux. Ainsi, le rapport
12C/13C passe de 88,99 pour les carbonates minéraux de référence à des valeurs
comprises entre 90,8 et 91,7 pour les molécules organiques biologiques. Ce
rapport est compris entre 90,2 et 92,4 pour la matière organique des sédiments
du Groenland. Ces valeurs suggèrent, mais ne prouvent pas de manière absolue,
l'existence d'une activité photosynthétique, donc d'une vie primitive, il y a
3,8 milliards d'années. En effet, cette matière organique très ancienne
(quelques fois réduite à des cristaux de graphite) a subi d'importantes
modifications au cours de la diagénèse. Le produit
final de cette dégradation, les kérogènes, se composent
de macromolécules complexes stables résistantes, qui peuvent même être
transformées en graphite pur pendant le métamorphisme. Tous ces traitements ont
très bien pu générer les enrichissements en 12C observés. Il faut aussi se
méfier terriblement de la contamination éventuelle de ces roches par des
microorganismes plus récents, contamination qui va, évidemment, biaiser les
analyses. A cause des multiples transformations subies par ces roches, il y a
fort peu de chances d'y retrouver des vestiges de microfossiles. Effectivement,
aucunes structures ressemblant à des bactéries fossiles n'ont été découvertes
dans les sédiments du Groënland.
Les microorganismes fossiles les plus anciens ont été trouvés dans
les sédiments de Barberton en Afrique du Sud et du Pilbara en Australie. Ces sédiments, vieux de 3,2 à 3,5
milliards d'années, sont légèrement plus jeunes que les roches du Groënland. Les sédiments sont bien conservés et montrent
l'existence d'une vie foisonnante dans des eaux littorales de faible
profondeur, et peut-être même proche de la surface de l'eau (certains biofilms présentent une structure lamellée qui semble
indiquer une vie bactérienne utilisant déjà l'énergie solaire). Les
microfossiles identifiés comprennent des structures filamenteuses longues de
dix à quelques centaines de microns de long, des bâtonnets de quelques microns
de long et des structures sphériques et ovoïdes d'environ 1 micron de diamètre.
Ces structures ont été attribuées à des bactéries fossilisées. La quantité de
carbone restant liée à ces microfossiles est généralement très faible (entre
0,01-0,5% avec des pointes exceptionnelles à 1%) ce qui rend l'analyse du
carbone organique particulièrement difficile. Les isotopes du carbone ont
cependant pu être mesurés et présentent un enrichissement variable mais
néanmoins significatif en carbone 12.
Arguant d'une ressemblance entre les cyanobactéries modernes et
les microfossiles du Pilbara, William Schopf, de l'université de Los
Angeles, a décrit ces derniers comme étant des fossiles de cyanobactéries. Ces
bactéries ancestrales auraient donc déjà pratiqué la photosynthèse oxygénée.
Interprétation très importante car elle ferait remonter la photosynthèse
oxygénée loin en arrière dans les temps géologiques alors que les indices
biochimiques les plus anciens de la photosynthèse oxygénée trouvés dans des
schistes carbonés, également en Australie, ne remontent qu'à 2,7 milliards
d'années. Selon l'Anglais Martin Brasier, de l'université d'Oxford, les
structures seraient bien constituées de matière carbonée organique enrichie en
carbone 12 (ce qui habituellement signe une origine biologique), mais la
matière organique serait d'origine purement chimique et non biologique. Elle
pourrait provenir de la réaction de l'hydrogène sur le monoxyde de carbone
(réaction dite de Fischer-Tropsch), deux gaz présents
dans les fluides des sources hydrothermales. L'accumulation de la matière
organique en microstructures serait due à la cristallisation du quartz dans la
veine hydrothermale, l'enrichissement important en carbone 12 résultant de
processus purement chimiques. L'explication de Brasier n'est cependant pas
totalement convaincante car on voit mal comment la réaction de Fischer-Tropsch pourrait produire des macromolécules aussi complexes
que les kérogènes (matière organique complexe, insoluble dans les solvants
usuels) déposés dans les veines hydrothermales. Une explication intermédiaire
pourrait être apportée par les travaux réalisés à Orléans, au Centre de
biophysique moléculaire du CNRS, par Frances Westall.
Des morphologies de microfossiles tels que biofilms,
polymères, coques, filaments, bâtonnets, ont été observés au microscope
électronique dans des échantillons de silice prélevés à Pilbara
dans des zones jouxtant les veines hydrothermales de Schopf,
mais jamais à l'intérieur même des veines hydrothermales. Ces morphologies
contiennent du carbone identifié par microanalyse au microscope électronique.
Il semble bien que les bactéries ancestrales vivaient, puis ont été fossilisées,
dans des roches sédimentaires au voisinage des veines hydrothermales. Les
veines hydrothermales ont très bien pu entraîner la matière organique des
bactéries mortes et/ou fossilisées (donc enrichis en carbone 12), matière
organique qui aurait été redéposée plus haut dans les veines hydrothermales,
pour former les fameuses structures carbonées complexes décrites par Schopf. Les structures de Schopf
ne seraient donc que des restes de matière organique bactérienne et non pas des
bactéries fossilisées. Cette explication est donc intermédiaire entre le tout
bactérien de Schopf et le tout chimique de Brasier.
Elle conforte néanmoins la présence de vie bactérienne il y a environ 3,5
milliards d'années.
Il faut hélas se rendre à l'évidence : l'espoir de retrouver
des petits automates chimiques fossilisés depuis 4 milliards d'années, ou même
des molécules organiques qui constituaient ces automates, est pratiquement nul.
En effet, quatre facteurs ont contribué à effacer leurs indices sur
Terre : l'histoire géologique mouvementée de la Terre (et en particulier
la tectonique de plaque), l'érosion due à la présence permanente d'eau liquide,
les effets délétères du rayonnement solaire en absence d'une couche d'ozone
protectrice pendant 2 milliards d'années et la vie elle-même qui produit
d'énormes quantités d'oxygène il y a 2 milliards d'années, un poison pour les
molécules organiques réduites. Les premières pages du livre de l'histoire de la
vie vont probablement rester blanches à tout jamais.
Un automate chimique ne pourra être facilement reconstitué en tube
à essais que s'il est simple. La datation des cratères d'impacts lunaires
suggère que les planètes du système solaire ont été soumises à un bombardement
intense il y a 4 milliards d'années au moment où la vie a probablement fait son
apparition sur Terre. Un petit nombre de ces impacts fut capable d'évaporer
toute l'eau des océans primitifs. De ce fait, les automates chimiques devaient
être suffisamment robustes pour pouvoir survivre aux impacts météoritiques et
cométaires et éventuellement redémarrer après les plus gros impacts. Il est raisonable de penser que pour être robuste, la vie
primitive devait être relativement simple et capable d'être répétée en
plusieurs exemplaires.
Apporter la preuve de la relative simplicité des automates
primitifs
La découverte d'autres exemplaires de vies sur d'autres corps
célestes conforterait la relative simplicité de l'origine de la vie en
apportant la preuve de son caractère répétitif. En identifiant à ce jour plus
de quatre-vingt molécules organiques dans le milieu interstellaire, et en
particulier le formaldéhyde et l'acide cyanhydrique, les radioastronomes ont
démontré que la chimie organique est universelle. Il ne reste plus maintenant
qu'à rechercher la présence permanente d'eau liquide. Présente en surface, elle
révèle l'existence d'une atmosphère qui permet l'apport en douceur des
molécules organiques par le biais des micrométéorites. Comme les molécules
organiques peuvent également se former dans les sources chaudes sous-marines,
tout océan extraterrestre présentant les signes d'une activité hydrothermale
constitue également un habitat biotique possible.
La planète Mars est l'objet d'une attention toute particulière.
Les lits d'écoulements asséchés observés par les missions martiennes Mariner 9,
Viking 1 et 2, Mars Pathfinder et Mars Global Surveyor indiquent clairement que Mars a connu dans sa
jeunesse des écoulements permanents de fluides. Ecoulement d'eau ? De
glaciers ? De neige carbonique ? La sonde martienne Odyssey placée en 2001 en orbite martienne par la Nasa
était émis par
le sol martien aprèsgéquipée d'un
spectromètre qui mesure les rayons
impact des rayons cosmiques. Ce spectromètre a détecté de très
importantes quantités d'atomes d'hydrogène présents en surface aux pôles mais
également à soixante centimètres dans le proche sous-sol martien jusqu'à 60° de
latitude nord et sud. La présence d'hydrogène signe la présence de glace d'eau,
permettant ainsi d'attribuer la formation des lits d'écoulement à la présence
d'eau liquide coulant jadis à la surface de Mars. La présence permanente d'eau
suppose une température constamment supérieure à 0°C, température atteinte
probablement grâce à l'existence d'une atmosphère dense générant un effet de
serre important. Grâce à cette atmosphère, la planète a pu accumuler des
micrométéorites à sa surface à l'instar de la Terre. En 1976, les deux sondes
Viking ne détectèrent ni molécules organiques ni vie à la surface de Mars sur
une profondeur de quelques centimètres. En fait, le sol martien semble
renfermer des oxydants puissants produits par le rayonnement solaire dans
l'atmosphère et/ou par des processus photochimiques au niveau du sol. La
présence d'oxydants exclut toute accumulation de molécules organiques à la
surface de la planète. Des calculs de simulation suggèrent que la diffusion des
oxydants dans le sous-sol ne devrait pas dépasser une profondeur de trois
mètres. L'absence de matière organique à la surface de Mars pourrait également
être due à des processus de dégradation directe par les UV solaires,
l'atmosphère martienne ne possédant pas de couche protectrice d'ozone. Les
sondes Viking n'ont pas trouvé de molécules organiques à la surface de Mars
mais on dispose en mai 2005 de 34 météorites qui sont généralement considérées
comme provenant de Mars. Ce sont les météorites SNC, dont la fameuse météorite
ALH 84001 présentée comme renfermant des nanobactéries
martiennes fossilisées. Cette affirmation est aujourd'hui de plus en plus
battue en brèche. Les soi-disant "microfossiles" se sont très
probablement formés par des processus minéraux consécutifs à des infitration d'eau terrestre. La présence de cristaux de
magnétite présentée comme preuve de la présence de vie martienne dans la
météorite n'est pas non plus convaincante dans la mesure où des cristaux
identiques ont été obtenus en laboratoire, d'une manière non biologique.
Cependant, certaines de ces météorites SNC renferment des molécules organiques.
Les ingrédients qui ont permis l'apparition de la vie sur Terre étaient donc rassemblées sur Mars. Il est dès lors tentant de penser
qu'une vie élémentaire utilisant les mêmes ingrédients que la vie terrestre ait
pu apparaître et se développer sur la planète rouge.
En 1997, le robot martien Sojourner a
analysé six sols et cinq roches autour du site d'atterrissage dans Ares Vallis. Les sols analysés sont tous identiques et très
proches des sols analysés par les sondes Viking. Les roches, analysées sur
quelques microns seulement, sont partiellement recouvertes de poussière du sol.
La composition des roches rappelle celle de certains granites terrestres et se
rapproche de celle de la croûte terrestre. Cependant, les résultats obtenus
sont insuffisants pour pouvoir trancher entre une origine volcanique et
sédimentaire. En 2004 et 2005, les deux rovers
américains, posés dans Gusev Crater
et Meridiani Planum,
respectivement, ont confirmé la présence passée d'eau liquide à la surface de
Mars. Plus particulièrement, Opportunity a révélé des
affleurements de roches sédimentaires renfermant de l'hématite et de la jarosite déposées à partir de solutions aqueuses.
Le 2 juin 2003, l'Agence spatiale européenne a lancé la mission
martienne Mars Express du pas de tir de Baïkonour. La mission a placé un
orbiteur muni de caméras stéréoscopiques, de différents spectromètres pour
analyser l'atmosphère et la composition du sol et d'un radar capable de
fouiller le sol sur plusieurs kilomètres pour y rechercher l'eau liquide et la
glace. Peu après sa mise en orbite en décembre 2003, le spectromètre Oméga de Mars
Express, piloté par Jean-Pierre Bibring, a confirmé
la présence de glace d'eau au niveau de la calotte polaire Sud et les dépôts de
sulfates. La caméra stéréoscopique HRSC à haute résolution de l'Allemand
Gerhard Neukum a mis en évidence des terrains volcaniques
et glaciaires bien plus jeunes que tout ce que l'on connaissait. Le
spectromètre PFS (Planetary Fourier spectrometer) de l'Italien Vittorio Formisano
a détecté du méthane et de la vapeur d'eau dans l'atmosphère de Mars à la
verticale d'Arabia Terra, un sol riche en glace comme
l'a montré Mars Odyssey. La teneur en méthane
varierait entre 10 et 30 parties par milliard en volume. Le méthane a aussi été
détecté depuis la Terre par les observatoires de Hawaï et du Chili. Comme la
durée de vie du méthane n'est que de 300 ans dans les conditions martiennes, il
faut envisager une production permanente de ce gaz. Origine géologique due à un
volcanisme résiduel ou origine biologique due à des bactéries
méthanogènes ? Pour pouvoir trancher entre ces deux options, il faudrait
disposer d'autres biomarqueurs gazeux comme
l'ammoniac, et/ou le formaldéhyde dont les durées de vie sont
très courtes sous le climat martien. Un atterrisseur, appelée Beagle 2 en
hommage au bateau de Charles Darwin, devait poser une station d'analyse
martienne d'environ dix kilos à la surface de Mars capablede
prélever des échantillons dans le proche sous-sol à l'aide d'une
"taupe" auto-enfouisseuse et dans les
roches de surface à l'aide d'un bras manipulateur. Le bras manipulateur devait
placer contre les roches de surface un broyeur/carottier, une caméra, un
microscope, un spectromètre à rayon X (donnant la composition en éléments
chimiques) et un spectromètre Mössbauer (donnant le degré d'oxydation du fer).
Les échantillons prélevés devaient être analysés par un spectromètre de masse
installé sur l'atterrisseur après combustion par pallier de températures,
technique permettant d'analyser les molécules organiques en déterminant leur
nature, minérale ou organique, ainsi que les distributions isotopiques. La
sonde Beagle 2 s'est correctement séparée de l'orbiteur Mars Express en
décembre 2003 mais n'a plus jamais donné signe de vie depuis cette date.
Si la vie est apparue sur Mars, la recherche de ses vestiges
fossilisés sera rendue plus facile que sur Terre par le fait que les sédiments
martiens anciens n'ont pas été détruits par des processus tectoniques,
contrairement aux sédiments terrestres.
Au-delà de Mars
Europe, le satellite de Jupiter, pourrait bien présenter des
environnements marins ressemblant aux sources sous-marines terrestres. Europe
est le plus petit des satellites de Jupiter avec un rayon légèrement inférieur
à celui de la Lune. Il orbite à une distance d'environ six cent mille
kilomètres de Jupiter, donc suffisamment près pour être réchauffé par l'effet
de marée dû au champ gravitationnel très important de la planète géante. En
1979 et 1980, la mission Voyager avait déjà photographié Europe et montré que
sa surface était recouverte par de la glace entaillée de profondes crevasses.
Depuis, le vaisseau spatial Galileo a fourni de très
belles images montrant, notamment des blocs de glace ayant pivoté sur
eux-mêmes. La surface présente peu de cratères d'impacts ce qui suggère un
remodelage continu de la surface par des phénomènes cryovolcaniques
ou tectoniques. Selon l'un des modèles de structure interne proposé, il y
aurait un océan d'eau liquide sous quelques dizaines de kilomètres de banquise,
la chaleur nécessaire au maintien de l'eau à l'état liquide étant apportée par
les fortes marées internes. La surface présente également des stries en forme
d'arcs, interprétées comme provenant d'un stress dû à des marées océaniques
sous-glaciaires. Des dépôts de sels ont été observés à la surface d'Europe par
spectroscopie dans le proche infrarouge, dépôts qui pourraient provenir de
remontées d'eau océanique salée. Enfin, Galileo a
enregistré un champ magnétique induit dans le champ magnétique de Jupiter
traduisant la présence d'un conducteur électrique. L'océan d'eau salée pourrait
très bien jouer ce rôle de conducteur électrique. Toutes ces observations
plaident en faveur de l'existence d'un océan sous-glaciaire d'eau salée. Il est
maintenant important de savoir s'il existe sur Europe un magma capable de
transférer la chaleur du coeur planétaire vers le fond océanique pour créer des
sources hydrothermales et, par conséquent, des molécules organiques. L'énergie
de marée induite par Jupiter est-elle suffisante pour fondre les silicates à
des températures supérieures à 1200°C ? La mise en évidence d'un magma sur
Europe fait partie des objectifs prioritaires de l'exploration d'Europe
actuellement à l'étude. Si Europe a maintenu une activité de marée et une
activité hydrothermale sous-glaciaire, une vie bactérienne a pu y apparaître et
y est peut-être encore active aujourd'hui. Europe apparaît de plus en plus
comme un lieu privilégié du système solaire pouvant héberger de l'eau liquide
et une vie bactérienne en activité.
Le cas particulier de Titan
Titan, le plus gros satellite de Saturne, possède une atmosphère
dense de 1,5 bar constituée essentiellement d'azote (plus de 90%) mais aussi de
méthane et d'un peu d'hydrogène. L'atmosphère renferme également d'épais
brouillards d'aérosols organiques. Les observations glanées par la mission
Voyager et les mesures faites à partir de la Terre indiquent clairement la
présence de nombreux hydrocarbures et de nitriles dans ce milieu. Parmi ces
composés organiques figurent l'acide cyanhydrique, l'acétylène, le cyanoacétylène, véritables passages obligés de la chimie prébiotique. La modélisation des processus physicochimiques
supposés se dérouler à la surface de Titan suggère la présence d'océan(s) de
méthane et d'éthane liquide en équilibre avec les constituants de l'atmosphère.
Titan représente un véritable laboratoire de production de composés prébiotiques à l'échelle planétaire. Bien que des traces de
vapeur d'eau aient été récemment détectées par le satellite ISO dans la haute
atmosphère, la température très basse (-180°C) régnant près de la surface
interdit la présence d'eau liquide. Les molécules ne peuvent donc pas évoluer
vers une vie de type terrestre. La mission NASA-ESA Cassini-Huygens
lancée en octobre 1997 a déposé la sonde Huygens à la surface de Titan le 14
janvier 2005 après plus de 7 années d'un voyage interplanétaire d'environ 1,5
milliards de kilomètres. La sonde a photographié des lits d'écoulements
asséchés façonnés vraisemblablement par du méthane liquide. Vers quels systèmes
complexes évoluent ces molécules en l'absence d'eau ? Existe-t-il une vie
exotique basée sur la chimie du carbone et le méthane liquide ? Les
instruments de Huygens ont analysé les molécules organiques pendant 150
minutes. Malheureusement, la sonde Huygens ne portait aucun instrument capable
de mesurer la chiralité de ces molécules organiques, véritable ligne de
démarcation entre matière inerte et matière vivante.
Une vie extra-solaire
La chasse aux planètes extrasolaires se pratique habituellement en
mesurant la perturbation apportée à l'étoile par la planète. En effet, lorsqu'une
planète gravite autour d'une étoile, l'étoile elle-même se met à valser autour
du centre de gravité commun étoile/planète. Plus la planète est massive et
proche de l'étoile, plus la vitesse de rotation de l'étoile est grande. C'est
ainsi que la vitesse du Soleil induite par Jupiter est de treize mètres par
seconde tandis que celle induite par la Terre n'est que de dix centimètres par
seconde. On peut rechercher une variation de la position de l'étoile dans le
ciel mais la précision des instruments de mesure actuels ne permet pas encore
une telle mesure. L'Agence spatiale européenne développe actuellement le projet
interférométrique Gaia qui devrait détecter les
planètes depuis l'espace. Gaia doit cartographier plus d'un milliard d'étoiles
de notre galaxie, y compris les étoiles les plus proches du Soleil. On peut
également observer des petites variations dans l'éloignement de l'étoile par
rapport à l'observateur. Lorsqu'une étoile accompagnée d'une planète décrit un
mouvement autour du centre de gravité commun, l'étoile s'éloigne et se
rapproche périodiquement de l'observateur et la lumière qu'elle émet
varie : la lumière est déplacée vers le bleu lorsque l'étoile se rapproche
de l'observateur et vers le rouge lorsqu'elle s'en éloigne. La meilleure précision
obtenue avec les instruments actuels est de quelques mètres par seconde, soit
une planète de la taille de Jupiter ou supérieure à celle-ci. C'est la méthode
qu'utilisèrent Michel Mayor et Didier Queloz à l'Observatoire de Haute-Provence.
En septembre 1995, après un suivi systématique des vitesses d'une centaine
d'étoiles, ils découvrirent la présence d'un corps de la moitié de Jupiter en
orbite autour de l'étoile 51 Pegase. L'objet
découvert était très proche de l'étoile et avait une température de 1500°C.
Quatre mois plus tard, Marcy et Butler décrivaient
deux objets ayant 2,8 et 6,4 fois la masse de Jupiter près des étoiles 70
Virgin dans la constellation de la Vierge et 47 Uma
dans la constellation de la Grande Ours. Les objets sont situés plus loin de
l'étoile. La planète 70 Virgin, géante et probablement gazeuse, est peu propice
à la vie. Elle peut, cependant, à l'instar de Jupiter et Saturne, avoir des
satellites de la taille de la Terre avec des températures permettant la
présence d'eau liquide. Fin mai 2005, le catalogue comptait 155 planètes
géantes extrasolaires, la plus petite d'entre elles
représentant 14 fois la Terre. Enfin, la méthode dite des "transits"
consiste à détecter l'ombre d'une planète lorsqu'elle passe devant son étoile
et provoque une mini-éclipse. Il s'agit donc de
mesurer la faible occultation passagère de l'étoile provoquée par le passage de
la planète devant celle-ci. En mars 2005, cinq exoplanètes
avaient pu être mises en évidence à l'aide de cette méthode. La mission française
COROT, en préparation, a précisément pour objectif de détecter des planètes
extrasolaires selon cette méthode, en plus des exoplanètes
géantes détectable par la méthode des perturbations décrite plus haut. La
variation de luminosité sera directement donnée par le rapport entre la surface
apparente de la planète et celle de l'étoile, soit environ 0,01 % pour une
planète de la taille de la Terre autour d'une étoile comme le Soleil., COROT
devrait donc pouvoir détecter des planètes telluriques en orbite autour
d'autres étoiles que notre Soleil. De plus COROT sera en mesure d'étudier la
composition des atmosphères, et donc de détecter des planètes semblables à la
Terre ou à Vénus.
La recherche de la vie sur les planètes extrasolaires ne peut se
faire que par l'analyse spectrale de ses manifestations, singularités dans
l'atmosphère et/ou message électromagnétique "intelligent" ([Seti], Search for Extraterrestrial
Intelligences). L'atmosphère terrestre renferme en permanence 21% d'oxygène
alors que les atmosphères des autres planètes du système solaire n'en
renferment que des traces. L'oxygène dans l'atmosphère terrestre est une
singularité à deux titres : il est surabondant par rapport à la croûte
terrestre et il devrait normalement disparaître par recombinaison avec les
roches. Sa présence permanente est, bien entendu, liée à l'existence d'une vie
intense à la surface de la terre et ne manquerait pas d'attirer l'attention de
tout observateur extraterrestre à la recherche de la vie. La présence de
grandes quantités d'oxygène atmosphérique va être révélée par la raie
caractéristique de l'oxygène à 760 nm à l'aide d'un spectrophotomètre dans la spectre visible de la planète. Pour des raisons
pratiques, il est plus facile de rechercher la signature de l'ozone O3, dans le
m. Dans l'hypothèse, extrêmement séduisante, où demspectre infrarouge à 9,6 l'oxygène atmosphérique extraterrestre serait
mis en évidence, les sceptiques ne manqueraient pas de faire remarquer que
l'oxygène peut être produit par des mécanismes chimiques non biologiques. Quoi
qu'il en soit, la présence simultanée d'ozone (donc d'oxygène), de vapeur d'eau
et de dioxyde de carbone apparaît aujourd'hui comme une signature probante
d'une vie planétaire exploitant largement la photosynthèse. Vers 2020, deux
projets seront dédiés à la recherche d'exoplanètes de
type terrestre. Le projet américain TPF (Terrestrial Planet Finder) et le projet européen Darwin/IRSI (Infrared Space Interferometer). Ce dernier consiste à placer une flotille de six télescopes spatiaux qui seront couplés dans
l'espace pour analyser les atmosphères planétaires par interférométrie et y
rechercher des singularités dues à une activité biologique. Enfin, la détection
par radioastronomie d'un signal électromagnétique intelligent, signal
provenant d'une civilisation extraterrestre ayant atteint un niveau suffisant
de développement technologique, apporterait la preuve indéniable de l'existence
d'une vie extraterrestre. Le programme d'étude mérite d'être soutenu même si, a
priori, la probabilité pour qu'une vie bactérienne extrasolaire évolue vers des
systèmes vivants exploitant l'électromagnétisme ou la lumière laser soit
extrêmement faible.
Conclusion
L'année 2003 a marqué le cinquantième anniversaire de l'expérience
remarquable de Miller. En démontrant qu'il était possible de former des acides
aminés - les briques des protéines - à partir de méthane, le représentant le
plus simple des molécules organiques, Miller suscita un énorme espoir :
les chimistes allaient peut-être pouvoir reconstituer une vie primitive en tube
à essais. Force est de constater que le rêve n'a pas encore été réalisé.
Pendant de très nombreuses années, les chimistes se sont efforcés de
reconstituer en laboratoire les trois familles de molécules indispensables au fonctionnement
de la cellule : les molécules de l'information ARN et ADN, les molécules
catalytiques et les molécules de compartimentation. Ils ont réussi à
reconstituer des précurseurs de membranes et des mini-protéines.
Par contre, les chimistes n'ont pas réussi à reconstituer les molécules d'ARN.
Même si la vie cellulaire a été précédée par un monde vivant d'ARN capable de
fournir à la fois le plan de montage et l'outil pour effectuer
l'autoreproduction, le problème reste entier car la synthèse des ARN primitifs
reste encore incomprise.
Les chances de succès vont dépendre de la simplicité des automates
à construire. Aussi est-il important de découvrir un deuxième exemple de vie
qui conforterait l'idée de simplicité en démontrant le caractère ubiquitaire de
la vie. Ce nouveau rêve peut-il devenir réalité ? Les biologistes ont
montré que la vie bactérienne peut survivre en s'adaptant à des conditions
extrêmes (hautes et basses températures, hautes pressions, forte salinité) et à
des environnements très diversifiés (fonds océaniques, sous-sol profond,
pergélisol, espace), ce qui élargit considérablement l'éventail des niches
biologiques extraterrestres. Les planétologues et les astronomes nous ont fait
découvrir des habitats possibles dans le système solaire et même au-delà. A ce
jour, la vie n'est connue que sur Terre et nul n'a le droit de généraliser à
partir d'un seul exemple. C'est la raison pour laquelle la recherche d'une
"seconde vie" est devenue une priorité scientifique pour les années à
venir. Tant que l'on ne dispose que d'un seul exemplaire de vie, on ne peut pas
exclure l'idée que la vie terrestre résulte de la rencontre extraordinaire et
unique d'un très grand nombre de molécules. Si dans plusieurs décennies, aucun
autre exemple de vie n'a été découvert dans les nombreux habitats
extraterrestres répertoriés, aucun signal intelligent n'a été perçu, ni aucune
vie n'a pu être reconstituée en tube à essais, alors il faudra accepter l'idée
que la vie terrestre pourrait bien être unique, que nous sommes probablement
seuls dans le vaste Univers et que nous ne saurons probablement jamais comment
la matière devint vivante, voilà 4 milliards d'années sur une petite planète,
ni trop grosse, ni trop petite....
Pour en savoir plus :
Livres
L'évolution chimique et les origines de
la vie, Brack, A. et Raulin,
F., Masson (1991).
La chimie du vivant. De la protéine à la photosynthèse, Brack,
A. et Mathis, P., Quatre à Quatre, Le Pommier La vie
est-elle universelle ? Brack,A.
et Leclercq, B., EDP Sciences (2003).
Et la matière devint vivante, Brack, A. Le Collège de
la Cité, Editions Le Pommier(2004).
Chapitres d'ouvrages
Origine de la vie, Brack,
A., CD Rom, Encyclopaedia Universalis
(1997).
L'asymétrie du vivant, Brack, A., Pour la Science,
Dossier Hors-série Les symétries de la nature,
Juillet 1998, pp.36-43.
Origine de la vie, Brack, A., Supplément de l'Encyclopaedia Universalis,
pp.743-750 (1999).
Sommes-nous seuls ? Brack,A.,
in Le Grand Livre du Ciel, Bordas, pp. 92-115 (1999)
Origine et distribution possible de la vie dans l'Univers, Brack,
A., in Qu'est-ce que la vie ? Université de Tous les Savoirs, Odile Jacob,
pp.37-46 (2000).
De l'origine de la vie terrestre à la vie dans l'Univers, Brack,
A., in L'origine de la vie sur Terre et dans l'Univers : L'Environnement
de la Terre primitive, Eds. M. Gargaud, D. Despois et J.P. Parisot, Presses Universitaires de Bordeaux,
pp.17-28 (2001).
De la naissance de la vie sur Terre à une vie universelle, Brack,
A., in Sur les traces du vivant : de la Terre aux étoiles, Eds. F. Raulin-Cerceau, P. Lena
et J. Schneider, Le Pommier, pp. 71-95 (2002).
La chimie de l'origine de la vie, Brack, A., in
L'origine de la vie sur Terre et dans l'Univers : Les traces du vivant,
Ed. M. Gargaud, Presses Universitaires de Bordeaux (2003) pp. 61-81.